Оплодотворение*
Энциклопедия Брокгауза Ф.А. и Ефрона И.А. (1890 - 1916гг.) Статьи для написания рефератов, курсовых работ, научные статьи, биографии (118447 статей и 6000 рисунков).
|
|
| А | Б | В | Г | Д | Е | Ё | Ж | З | И | Й | К | Л | М | Н | О | П | Р | С | Т | У | Ф | Х | Ц | Ч | Ш | Щ | Ы | Э | Ю | Я | A | B | C | D | E | F | G | H | I | J | K | L | M | N | O | P | Q | R | S | T | U | V | W | Z |
| О | ОА | ОБ | ОВ | ОГ | ОД | ОЕ | ОЖ | ОЗ | ОИ | ОЙ | ОК | ОЛ | ОМ | ОН | ОО | ОП | ОР | ОС | ОТ | ОУ | ОФ | ОХ | ОЦ | ОЧ | ОШ | ОЩ | ОЭ | ОЯ |
| ОПА |
| ОПЕ |
| ОПЗ |
| ОПИ |
| ОПЛ |
| ОПО |
| ОПП |
| ОПР |
| ОПС |
| ОПТ |
| ОПУ |
| ОПЫ |
| ОПЬ |
Оплодотворение* — Оплодотворение* в широком смысле слова называется слияние клеточек, животных или растительных, после которого они получают или восстановляют способность нормально размножаться делением. В этом смысле слова под Оплодотворение* понимаются как процессы типического Оплодотворение*, свойственного многоклеточным животным, а также семянным и многим споровым растениям, так и процессы конъюгации и копуляции одноклеточных организмов и некоторых многоклеточных растений. Оплодотворение* в тесном смысле слова состоит в соединении двух клеточек, яйцевой и семянной, причем протоплазма сливается с протоплазмой, ядро с ядром и центрозома (см. Клеточка) с центрозомой, так что при последующем делении оплодотворенной лицевой клеточки новые клеточки получают составные части обеих слившихся. Процесс Оплодотворение* в той или иной форме широко распространен в органическом мире, и хотя известны некоторые организмы, обладающие, по-видимому, способностью безгранично размножаться без Оплодотворение*, но правилом мы должны считать периодически наступающую необходимость слияния между собой клеточек, полного или временного и частичного, без которого дальнейшее размножение этих организмов или становится невозможным, или идет ненормально и скоро прекращается окончательно. Значение процесса Оплодотворение* в органическом мире очевидно уже по громадной массе таких анатомических, физиологических и биологических особенностей различных организмов, которые обеспечивают возможность Оплодотворение* Необходимость для одноклеточных организмов Оплодотворение* (которое здесь совершается в виде конъюгации или копуляции) с особенной очевидностью доказана исследованиями Мопа (Mopas) над ресничными инфузориями. Оплодотворенная путем конъюгации особь этих животных размножается путем деления, но после известного числа поколений, большего или меньшего, смотря по виду инфузории, а отчасти и по внешним условиям, между инфузориями данной культуры возникает потребность конъюгации, наступает "эпидемия конъюгации", причем почти все индивиды данной культуры оказываются в состоянии конъюгации. У Leucophrys patula потребность конъюгации обнаруживается лишь после 300 поколения, происшедшего путем простого деления, у Onychodromus grandis — после 140, у Stylonychia pustulata — после 130. Если же конъюгации в течение известного периода не произойдет, то наступают явления, которые Мопа назвал "старческим вырождением"; способность деления сохраняется, но получаются индивиды ненормальные, которые через несколько поколений вымирают, причем даже запоздавшая конъюгация не спасает их. У некоторых простейших организмов бесполое размножение правильно чередуется с конъюгацией, после которой снова начинается такой же цикл развития. При этом способность размножаться может в колониальных организмах быть свойственна каждой клеточке, в других случаях (напр. у Volvox) лишь некоторым клеточкам колонии. У некоторых групп высших животных единственным способом размножения является размножение с помощью оплодотворенных яиц (напр. у позвоночных, моллюсков); но во многих группах животных и растений наряду с размножением с помощью клеточек, нуждающихся в Оплодотворение*, происходит и размножение без Оплодотворение*, причем или развиваются клеточки, соответствующие нормальным яйцевым, но не нуждающиеся в Оплодотворение* (напр. партеногенетически развивающиеся яйца, споры), или же от организма отделяются целые группы клеточек (почки), или, наконец, весь организм подвергается процессу деления. В этих случаях размножение с помощью оплодотворяемых яиц чередуется с другими формами размножения полового (партеногенетические яйца) или бесполого (почкование, деление), причем размножение с Оплодотворение* может или чередоваться с размножением без Оплодотворение* совершенно правильно, или же такой правильности нет и число поколений, размножающихся без Оплодотворение*, может быть неопределенно большим; наконец, иногда из яиц одного поколения некоторые оплодотворяются, другие нет, хотя и те, и другие подвергаются дальнейшему развитию. Процесс Оплодотворение* в типических его формах лучше всего изучен на яйцах некоторых животных (иглокожих и Ascaris) и на зародышевом мешке семяпочек некоторых растений. Процессу Оплодотворение* у иглокожих предшествует выделение направляющих или путеводных телец (см. Направительные тельца); по выделении их яйца, окруженные студенистой проницаемой для сперматозоидов (живчиков, см.) оболочкой, выводятся из тела матери. Сперматозоиды (фиг. 1) проникают в студенистую оболочку и — навстречу тому из них, который проник глубже всего, образуется на поверхности яйца выступ; с этим выступом сперматозоид сливается и вместе с ним втягивается внутрь яйца, которое одевается на поверхности новой оболочкой (желточной); после образования последней проникновение внутрь яйца новых сперматозоидов становится невозможным.
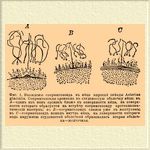
Фиг. 1. Вхождение сперматозоида в яйцо морской звезды Asterias glacialis. Сперматозоиды проникли в студенистую оболочку яйца; в А — один из них проник ближе к поверхности яйца, на поверхности которого образуется навстречу сперматозоиду протоплазматический выступ; в В — сперматозоид слился уже с выступом; в С — сперматозоид вошел внутрь яйца, на поверхности которого наружной студенистой оболочкой образовалась вторая оболочка — желточная.
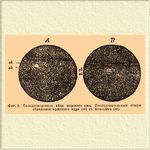
Фиг. 2. Оплодотворенное яйцо морского ежа. Последовательные стадии сближения мужского ядра (sk) с женским (bk).
Вхождением сперматозоида оканчивается внешняя часть процесса Оплодотворение* Головка сперматозоида, принимая в себя жидкость из протоплазмы яйца, превращается в небольшой пузырек (мужское ядро), окруженный светлым полем, который и движется внутрь яйца по направлению к ядру яйцевой клеточки (женскому ядру). Как около мужского, так и около женского ядра можно заметить при этом по особому маленькому тельцу (центрозом, центральному тельцу — см. Клеточка). Протоплазма вокруг ядер получает лучистое строение. Мужское и женское ядро сближаются между собой и, наконец, сливаются в одно ядро яйца, которое и подвергается затем делению при процессе дробления. Центрозомы делятся каждая на две части и части эти сливаются между собой так, что половина мужской центрозомы сливается с половиной женской; таким образом, получаются две центрозомы, которые и являются центрами при последующем делении яйца. Процессом слияния центрозом и заканчивается Оплодотворение* Особенности в Оплодотворение* яйца Ascaris megalocephala сводятся, главным образом, к следующему: образование направительных пузырьков происходит после вхождения в яйцо сперматозоида; мужское ядро проходит в центр яйца и к нему затем движется по окончании процесса выделения направляющих пузырьков женское ядро; слияния мужского ядра с женским не происходит непосредственно по их сближении; лишь когда яйцо подготовляется к первому делению, половина ядерного вещества мужского ядра сливается с половиной вещества женского, давая ядра двух первых бластомер (см. Дробление). Процесс Оплодотворение* у цветковых растений не отличается существенно от описанного процесса Оплодотворение* у животных. Роль яйца играет яйцевая клеточка зародышевого мешка, роль сперматозоида — клеточка цветневой трубочки; содержимое цветневой трубочки проходит в зародышевый мешок и ядра мужское и женское, а равно и центрозомы (по 2 при каждом ядре) сливаются между собой. В случаях, хорошо изученных, можно убедиться, что количество ядерного вещества в каждом из сливающихся ядер уменьшено вдвое по сравнению с обыкновенными ядрами клеточек тех же организмов, притом количество ядерного вещества в мужском и женском ядре (как можно убедиться, следя за изменениями в распределении хроматина) одинаково и вообще оба ядра должны считаться эквивалентными друг другу. Что касается роли частей сливающихся половых клеточек, то в этом отношении существует разногласие: одни считают слияние ядер, центрозом и протоплазм обеих клеточек за процессы в равной степени важные и необходимые, другие придают слиянию протоплазм лишь второстепенное значение, считая существенным лишь слияние ядер и центрозом. Эквивалентности яйца и мужской половой клеточки и одинаковое содержание в них ядерного вещества (различие в величине между мужским и женским ядром зависит от количества содержащихся в них жидких частей) наглядно обнаруживается, по Гертвигу, в развитии яиц и живчиков лошадиной аскариды до момента Оплодотворение* В первичных половых клеточках как самцов, так и самок этой аскариды ядро содержит по 4 хроматиновых нити, затем число их удваивается. Далее у самцов клеточка подвергается двум последовательным делениям, причем удвоения числа хроматиновых нитей не происходит и ядро каждой из 4 образующихся таким образом семянных клеточек, из которых каждая развивается в сперматозоид, содержит по 2 хроматиновых нити. В женских половых клетках после стадии с 8 хроматиновыми нитями наступает выделение направляющих пузырьков (т. е. в сущности деление лицевой клеточки с той лишь особенностью, что при одном из новых ядер отделяется лишь незначительная часть протоплазмы).
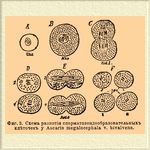
Фиг. 3. Схема развития сперматозоидообразовательных клеточек у Ascaric megalocephala v. bivalvens.
При образовании первого пузырька ядро яйцевой клеточки теряет 4 хроматиновых нити, при выделении 2-го — еще 2, и следовательно, яйцо, готовое к Оплодотворение*, заключает в ядре 2 хроматиновых нити, как и сперматозоид. Что касается направительных пузырьков, то первый из них, содержащий 4 хроматиновых нити, делится на 2 (каждый с 2 хроматиновыми нитями).
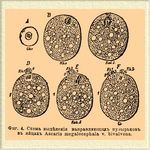
Фиг. 4. Схема выделения направляющих пузырьков в яйцах Ascaris megalocephala v. bivalvens.
Таким образом, как первичная мужская половая клетка, так и женские дают по 4 клеточки с половинным числом хроматиновых нитей, но между тем как все 4 мужские клетчатки способны к Оплодотворение*, из женских оказывается способной к Оплодотворение* лишь одна (яйцо), а три остальных (направляющие пузырьки) дальнейшего развития не получают и гибнут, представляя собой как бы недоразвитые, абортивные яйца. Способность половых элементов к Оплодотворение* продолжается обыкновенно более или менее короткое время, по прошествии которого способность эта утрачивается. В некоторых случаях половые клеточки сохраняют, однако, способность к Оплодотворение* сравнительно весьма долгое время: так семя, сохраняющееся в семеприемнике пчелы матки, весьма долгое время способно оплодотворять яйца, у некоторых млекопитающих, напр. у некоторых летучих мышей, козули — семя целые месяцы остается в теле самки и лишь после того оплодотворяет яйца. При Оплодотворение* в яйцо проникает обыкновенно лишь один сперматозоид, которого ядро и сливается затем с ядром яйцевой клеточки. В некоторых случаях наблюдается, однако, явление проникания внутрь яйца многих сперматозоидов — так называемая полиспермия. Различают полиспермию истинную, когда несколько мужских ядер сливаются с женским ядром, результатом чего является всегда неправильное и скоро останавливающееся дальнейшее развитие (при этом случается, что с женским ядром сливаются 2 мужских и яйцо начинает затем дробиться, но не на 2, 4 и т. д. бластомер, а сразу на 4, 8 и т. д.; иногда женское ядро при полиспермии распадается на несколько частей и каждая из них сливается с особым мужским ядром), и полиспермию ложную, когда из многих вошедших в яйцо сперматозоидов лишь один сливается с женским ядром (не слившиеся с женским мужские ядра могут при этом подвергаться делению в протоплазме яйца). Утверждают, что ложная полиспермия у некоторых животных (земноводных) явление весьма обыкновенное. Описанному процессу Оплодотворение* у высших животных и растений соответствует конъюгация и копуляция одноклеточных животных и многих как одноклеточных, так и многоклеточных растений. Особенное сходство с явлениями Оплодотворение* у высших животных представляет конъюгация ресничных инфузорий. Конъюгирующие индивиды прикладываются друг к другу частью поверхности тела и здесь между телами обеих особей устанавливается непосредственная связь. Как известно, у ресничных инфузорий наблюдается два рода ядер: большое главное ядро, макронуклеус (Macronucleus, Hauptkern немецких авторов) и меньшее (одно или несколько) придаточное или замещающее ядро, микронуклеус (Micronucleus, Nebenkern, Ersatzkern). При конъюгации первое из них распадается на части, и части эти растворяются или выбрасываются из тела — словом, главное ядро погибает. Придаточные ядра подвергаются процессу деления, часть получившихся таким образом ядер погибает подобно главному ядру, часть остается в теле животного, часть переходит в тело другой из конъюгирующих особей. Ядра, переходящие в другую особь, сливаются в ней с остающимися там, получается новое ядро из слившихся частей ядер обеих особей и затем такое ядро снова делится, давая новое главное и придаточное ядро. У Paramaecium canudatum (ф. 5) конъюгирующие особи прикладываются друг другу ротовыми поверхностями и здесь соединяются между собой, главное ядро начинает разрушаться, придаточное делится на 2, потом на 4 части, из которых 3 в каждом неделимом разрушаются (сходство с образованием направляющих пузырьков), остающееся делится еще раз (при чем происходит, по исследованиям Р. Гертвига, такое же уменьшение вдвое числа хроматиновых элементов, как в яйцах перед Оплодотворение*); одно ядро (стационерное) остается в теле особи, другое (блуждающее) переходит в тело другой, где и сливается со стационарным ядром ее.
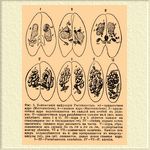
Фиг. 5. Конъюгация инфузории Paramaecium. nk — придаточное ядро (Micronucleus), k — главное ядро (Macronucleus). I — придаточное ядро подготовляется в каждой из особей к делению. II — придаточные ядра разделяются каждое на 4 (из них уцелеют лишь 1 и 5). III — ядра 1 и 5 делятся каждое на стационарное (w) и блуждающее (m). IV — обмен ядрами почти закончен; главное ядро распалось на части. V — ядра, образовавшиеся из слияния 5m c 1w и 5w с 1m, подвергаются новому делению. VI и VII — конъюгация окончена. Каждое из ядер подразделяется на 4: два превращаются в микронуклеусы (nk), два (pt) сливаются, давая новое главное ядро. I — III — Paramaecium caudatum, IV — VI — P. aurelia.
После этого обмена ядрами (которые в противоположность стационарным женским можно считать мужскими) и слияния их обе особи оказываются как бы взаимнооплодотворенными. Деление ядер, происшедших путем слияния, ведет к образованию в каждой особи главного и придаточного (или придаточных) ядер, а конъюгировавшие особи после обмена ядер разделяются. У некоторых сидячих ресничных инфузорий, напр. сувоек Vorticella, y Epistylis, наблюдается несколько иной процесс: часть особей делится, образовавшиеся мелкие особи отделяются от ножек и конъюгируют с крупными; внутри их происходят те же явления, как и у других инфузорий, но малая особь не получает способности дальнейшего развития и в конце концов вполне сливается с крупной. Здесь мелкие особи очевидно соответствуют мужским элементам (более подвижным и сливающимся с женскими, которые одни сохраняют способность дальнейшего развития). Конъюгация в различных группах животных и растений может совершаться весьма различно: иногда между сливающимися особями (двумя или несколькими — напр. у некоторых корненожек) не наблюдается никакой разницы. В других случаях для конъюгации (или копуляции) служат особые подвижные клеточки, снабженные мерцательными жгутиками (такие подвижные клеточки свойственны и животным и весьма многим водорослям); эти клеточки могут быть одинаковы или разной величины; наконец, во многих растениях лишь одна из сливающихся половых клеточек подвижна (мужская), другая же (женская) лишена каких-либо органов движения. Как показывает особенно конъюгация у низших животных и растений, которую мы бесспорно должны считать за простейший, наиболее примитивный акт Оплодотворение*, между мужскими и женскими оплодотворяющими и оплодотворяемыми половыми элементами не существует никакого коренного специфического различия; если же у многоклеточных животных и многих растений половые продукты представляют резкое различие по своему виду, величине и строению, то различие это является лишь результатом разделения труда между половыми клеточками: одни получили форму элементов весьма подвижных (активно или пассивно), другие отличаются обильным отложением запасных питательных веществ, относительно велики и неподвижны. Первые и являются мужскими элементами, вторые — женскими.
Н. Книпович.
|
Смотрии так же... |
|
|